apuntes pdf- Replicación, Transcripción, Traducción
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: REPLICACIÓN, TRANSCRIPCIÓN, TRADUCCIÓN-1/4
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: REPLICACIÓN-2/4
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: TRANSCRIPCIÓN-3/4
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: CÓDIGO GENÉTICO, TRADUCCIÓN-4/4
martes, 26 de abril de 2016
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: CÓDIGO GENÉTICO, TRADUCCIÓN-4/4

CÓDIGO GENÉTICO- TRADUCCION
Para iniciar este proceso es necesario conocer el código genético, es decir
la relación que existe entre la secuencia de nucleótidos y los correspondientes
aminoácidos.
Esta relación fue establecida a base de crear secuencias artificiales y
comprobar que aminoácido les correspondía. Así fue establecido el código
genético, tras los experimentos de Severo Ochoa, Nirenberg, Matthaei Korenberg y
Khorana.
Tras muchas pruebas llegaron a la conclusión de que cada tres nucleótidos
codificaban para un aminoácido. Esto supone que existen más tripletes que
aminoácidos, 64 tripletes, frente a 20 aminoácidos, así que la pregunta es
saber que ocurre con el “exceso” de tripletes.
De este modo se observó que los tripletes no solo codificaban para los
20 aminoácidos, sino que además existen tripletes sin sentido, es decir que no
codifican para ningún aminoácido, pero marcan el final de la codificación, es
decir, son secuencias stop.
Por otra parte se observa que diferentes tripletes codifican para el mismo
aminoácido.
De este modo se establecen las características generales del código
genético:
1- Es universal, es el mismo para todos los
seres vivos, lo que demuestra el origen común de todos los organismos
(incluidos virus).
2- Es degenerado, varios tripletes codifican
para el mismo aminoácido
3- Es una secuencia lineal
4- No tiene huecos, es decir, la secuencia de
nucleótidos es continua, codificando los aminoácidos de modo igualmente
continuo.
El ADN contiene la información que es transcrita al ARNm en forma de
tripletes o codones de ARN, éstos a su vez se corresponden con la secuencia
anticodón (triplete complementario) que lleva el ARNt.

Mecanismo de la
traducción
Primero se tienen que activar los
aminoácidos, esto ocurre cuando se unen al ARNt, proceso que requiere la
actuación de la aminoacil-ARNt-sintetasa, y ATP. El aminoácido que se une es el
codificado por el triplete complementario al anticodón.
Una vez unido comienzan las distintas fases:
1-Iniciación-
El ARNm se une a la subunidad menor del ribosoma,
por el extremo 5’
de la cadena. Para que se generen estas uniones interviene el IF3 (factor de
iniciación 3). Tras esto se une el ARNt que transporta el aminoácido metionina,
codificado por el triplete AUG (presente en el ARNm), triplete que marca el
inicio de la traducción de la secuencia del ADN. Para que esta unión sea
efectiva se requiere el IF2. Por último se une la subunidad mayor del ribosoma,
para ello es necesaria la actuación del IF1 y presencia de Mg+2. Cuando se ha
producido la unión de todas las estructuras se genera el complejo de
iniciación.
vídeo: activación aminoácidos
vídeos fases traducción (síntesis proteínas):
fase de Iniciación
fase de Elongación
fase de Terminación

2-Elongación-
El ribosoma se va desplazando a lo largo de la
cadena de ARNm, en sentido 5’-3’ .
El ribosoma
presenta una estructura tridimesional con 3 regiones o sitios, el sitio A, el sitio P y el sitio E,
cada uno ocupa el espacio equivalente a un triplete.
Una vez que ha entrado el primer ARNt con el aminoácido
(aa) met, y se acopla la subunidad mayor del ribosoma, el ARNt-aa queda ubicado
en el sitio P del ribosoma, quedando los sitio A y E libres. Al sitio A accede
otro ARNt, cargado con el aa correspondiente al triplete que aparece en el
ARNm. Una vez que se encuentran situados el
1º aa met y el 2º aa, se produce la unión entre ellos, por la acción de
la aminoacil-peptidasa (en realidad la propia acción catalítica del ribosoma,
es una ribozima). Una vez unidos ambos
aa, el 1º se libera de su correspondiente ARNt. El ribosoma se desplaza el
equivalente a un triplete, de modo que el primer ARNt se situa en el sitio E, y
el 2º ARNt en el sitio P, quedando libre el sitio A que permite la entrada de
un nuevo ARNt cargado con el aa correspondiente a la secuencia del ARNm. Igual
que antes se producirá la unión entre este aa y los dos anteriores. La
repetición de este proceso lleva a la elongación de la cadena, añadiendo los aa
de acuerdo con la secuencia indicada en el ARNm.
3-Terminación-
En el desplazamiento del ribosoma por el
ARNm termina apareciendo un triplete sin sentido, es decir, un triplete que
indica el término de la cadena (triplete stop), lo que da lugar a la liberación
de las subunidades del ribosoma y de la cadena proteica, con la colaboración de los factores de
liberación (RF).
vídeo: síntesis proteínas

Regulación de la
expresión genética
La célula tiene que controlar que cantidad de proteínas producir. En
procariotas Jacob y Monod realizaron un experimento (1960) en el que observaron
la existencia de un operón—conjunto
de genes que llevan distintas informaciones para regular como producir las
proteínas.
En este operón aparecen:
- genes estructurales- secuencia
de ADN que llevan la información para la producción de la proteína activa.
- genes promotores- secuencia de
ADN donde se una la ARN-polimerasa para la transcipción
- genes reguladores- genes que
llevan información para crear una proteína reguladora.
- genes operadores- donde se va
a unir la proteína reguladora y al
unirse bloquea la transcripción.
-Operón inducible-
El proceso catabólicos, en los que reciben sustancias complejas y que se
convierten en sustancias más sencillas, se produce un proceso inducible.
La secuencia está siempre bloqueada, es decir, el regulador ha producido la
proteína que bloquea al operador, de modo que impide que la ARN polimerasa
pueda leer. Esto se activa cuando aparece el sustrato a degradar. Este sustrato
se unirá a la proteína que bloquea al promotor, desbloqueando así la transcripción
y permitiendo que actúe la ARN polimerasa que realiza la transcripción
-Operón represible-
En procesos anabólicos en los que de moléculas sencillas se forma una
molécula compleja, se produce un efecto represor.
La transcripción está activa, se están formando proteínas, y cuando ya se
han producido suficientes, estas propias proteínas se une a la proteína
reguladora que se unirá al operador, bloqueando así la actividad de la
ARNpolimerasa, y por tanto la transcripción.
ANTERIOR apuntes pdf
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: TRANSCRIPCIÓN-3/4


TRANSCRIPCION
Características generales
Mecanismo por el que se inicia un proceso de copia de un fragmento de ADN
para finalmente poder producir una proteína.
De la cadena de ADN se realiza una copia, pero de ARN (por lo que la timina
será sustituida por uracilo) e
intervendrán ARN polimerasas. En
eucariotas en el ADN se presentan secuencias no codificadoras (intrones) y otras codificadoras (exones), estructuras que no se
presentan en los procariotas.
En eucariotas se genera un transcrito primario que debe sufrir un proceso
de maduración para eliminar los intrones y que se unan los exones, pero en
procariotas, aunque también presentan un procesamiento, no existe eliminación
de intrones.
Por otra parte en procariotas todo el proceso ocurre en el citoplasma,
mientras que en eucariotas la copia y procesamiento del ARN tienen lugar en el
núcleo y de éste sale el ARN maduro hacia el citoplasma.
Los procariotas son además policistrónicos,
es decir la misma secuencia de ARNm genera, codifica para distintas proteínas,
por la modificación en el patrón de lectura, además de llevar varias secuencias
codificantes. Los eucariotas son monocistrónicos,
es decir, tras el procesamiento y eliminación de los intrones y unión de los
exones, queda una única secuencia codificadora de modo que se genera una única
proteína.
vídeo: transcripción básica
transcripción procariota
transcripción/traducción-comparación procariotas/eucariotas

Etapas de la
transcripción: (semejantes en procariotas y eucariotas)
1-
Iniciación
La enzima ARNpolimerasa se une a una secuencia de
bases nitrogenadas de la cadena de ADN que reconoce (centro promotor, TATA box),
en procariotas son necesarios factores de reconocimiento “rho y sigma”. Se abre
la doble hélice, la síntesis siempre se realiza en sentido 5’-3’los nucleótidos
que van a ser añadidos son nucleótidos trifosfato, son ellos los que llevan la
energía para la unión
2-
Elongación
Se añaden los nucleótidos trifosfato en sentido
5’-3’ , de
ello se encarga la ARNpolimerasa I, que en el caso de procariotas forma un
única secuencia de ARN a partir de la cual se formarán todos lo tipos
diferentes de ARNs.
En el caso de eucariotas existen distintos tipos
de ARN polimerasas:
ARNpolimerasaI- se encarga de las copias de ARNr
ARNpolimerasaII-se encarga de las copias de ARNm
ARNpolimerasaIII- se encarga de las copias de ARNt
Por otra parte en eucariotas una vez comenzada la
elongación se añade en el extremo 5’
una caperuza de metilguanosina trifosfato, para que este ARN no sea dañado o
atacado por otras enzimas.
3-
Terminación
El proceso de elongación acaba cuando la
ARNpolimerasa reconoce una secuencia en la que indica que el fragmento de debe
ser copiado finaliza ahí. En el caso de procariotas son secuencias
palindrómicas.
En el caso de eucariotas una vez
reconocida la secuencia de terminación se añade una cola poliA en el extermo 3’ , unos 200-300 nucleotidos de
A, para proteger toda la cadena.


Proceso de Maduración
Tanto en procariotas como en eucariotas existe un proceso de maduración de
esta secuencia se ARN recién transcrita, llamada ARN transcrito primario. Pero en el caso de procariotas no es tan
manifiesta como en eucariotas. En éstos últimos este proceso de maduración
recibe el nombre de splicing, que
consiste en la eliminación de las secuencias no codificantes (los intrones) y
la unión de las codificantes (los exones). Ese proceso se lleva a cabo en el
interior del núcleo a través de un complejo enzimatico el espliceosoma, que rompe la cadena de ARN en el punto del intrones y
une los exones.
vídeo: espliceosoma: eliminación de intrones

ANTERIOR apuntes pdf SIGUIENTE
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: REPLICACIÓN-2/4
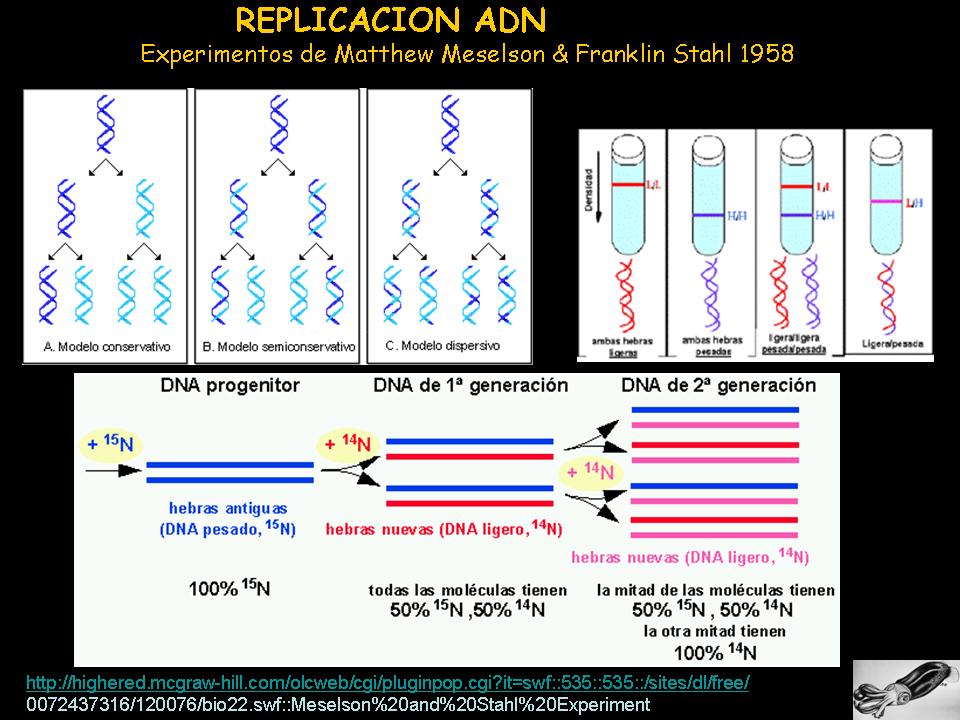
Meselson y Stahl (1958) intentaron averiguar como se copiaba (replicación) el
ADN. Llegaron a la conclusión de que el proceso era semiconservativo. Para demostrarlo prepararon unas muestras de ADN
con N pesado (15N), que posteriormente fue introducido en un medio con N normal
(14N). En el medio con N15 las bacterias lo incorporan en su material genético,
de modo que éste será más pesado. Cuando son transferidas a un medio con N14
seré éste el que se incorpore, variando así el peso de la molécula. Tas los
cultivos se sometieron a centrifugación, para aislar el ADN, y observar su peso
a través de las diferentes bandas que se pudieran formar
Las opciones que propusieron en cuanto a como se copiaba (replicaba) el
material genético fueron:
-
hipótesis conservativa- la cadena de ADN sirve de molde para
formar una nueva, pero las cadenas viejas y las nuevas quedan por separado
formando las dobles hélices, es decir, una doble hélice lleva las 2 cadenas
viejas (pesadas, N15) y la otra doble hélice las 2 cadenas nuevas (ligeras
N14), lo que mostraría 2 claras bandas de acuerdo a su peso, banda pesada y
banda ligera.
-
hipótesis dispersiva- la cadena de ADN sirve de molde, pero
las dobles hélices se forman a partir de fragmentos tanto de las cadenas viejas
como de las nuevas. Esto originaría una mezcla de ambos pesos, lo que no daría
lugar a una banda definida, se formaría una zona ancha correspondiente e
intermedia, correspondiente a la mezcla de ambos pesos N15-N14.
-
hipótesis semiconservativa- la cadena de ADN sirve de molde, pero
las dobles hélices se forman con una cadena vieja y otra nueva, así ambas
dobles hélices llevan tanto una cadena molde como otra de nueva síntesis. Tras dos
divisiones celulares se observará una banda de peso intermedio (hebra pesada
N15 + hebra ligera N14) y otra banda de peso ligero (hebra ligera N14 + hebra
ligera N14).
El experimento demostró que las nuevas dobles hélices portaban una cadena
con N15 y la otra con N14, es decir, una hebra antigua y otra nueva, de modo
que se confirmaba la hipótesis semiconservativa.
vídeo: experimento Meselson & Stahl- replicación semiconservativa
replicación semiconservativa

vídeo: Replicación bidireccional ADN-circular-procariota
REPLICACIÓN
La replicación es el mecanismo que permite la copia exacta de la información
contenida en el ADN. Este proceso tiene lugar para mantener la información que
será transferida a las células hijas (tanto en células procariotas como
eucariotas). Ocurre en ambos grupos celulares, aunque con ciertas diferencias.
CÉLULA PROCARIOTA-BACTERIAS
Las células procariotas tienen un cromosoma circular único de ADN. La copia
del material genético empieza en un
único punto, Ori C, que genera una burbuja de replicación (separación, apertura
de la doble cadena de ADN), replicón. En procariotas solo existe una burbuja de
replicación, mientras que en eucariotas hay multiples, al ser cromosomas más
largos.
El ADN está formado por dos cadenas antiparalelas (5’-3’/3’-5’ ), ambas cadenas son copiadas
y la copia es bidireccional (se copia en ambas direcciones). En uno de los
lados se copia al tiempo que se va abriendo la burbuja de replicación (la doble
hélice). El punto que se va abriendo para ir replicando recibe el nombre de horquilla de replicación. Para permitir
estos procesos (apertura de la doble hélice y copia) intervienen diversas
enzimas y proteínas:
-para separar ambas hebras interviene la enzima helicasa, a medida que se abre la horquilla aumenta la compactación
de la hebra, para evitarlo actúan las topoisomerasas
I, II (= girasa). Además para mantener la hebra abierta actúan las proteínas
estabilizadoras de la hebra las SSB.
A continuación actúa la ADN-polimerasa
III que añade los nucleótidos complementarios de la cadena que se está
copiando, pero estos nucleótidos solo pueden añadirse en sentido 5’-3’ , de modo que tiene que copiar
la cadena de sentido opuesto, la que presenta sentido 3’-5. Solo una de las
cadenas presenta el sentido que se necesita a la vez que se va abriendo, a esta
cadena se la llama hebra conductora.
Mientras que la otra hebra presenta sentido antiparalelo, se va a llamar hebra retardada. Esta hebra se va a
copiar por fragmentos (fragmentos de
Okazaki).
Por otra parte, la ADN-polimerasa III no puede actuar sin tener un apoyo
previo, para poder actuar necesita que ya exista un pequeño fragmento de nucleótidos.
Este fragmento lo coloca una ARN
polimerasa, sitúa un pequeño fragmento llamado primer
o cebador.
A medida que la horquilla de replicación se va abriendo, las dos hebras
(una en sentido 5’-3’
y la otra 3’-5’ )
se van copiando, pero la 3’-5’ será la hebra conductora, ya que permite la
adición de nucleótidos en sentido 5’-3’ ,
a la vez que la propia doble hélice se va abriendo. Esta cadena se copia de
forma continua y más rápida. La hebra
complementaria, la 5’-3’ (la hebra retardada),
muestra un sentido antiparalelo a la adición de nucleótidos, (adición siempre en
sentido 5’-3), de modo que ha de copiarse en sentido opuesto al de apertura de
la doble hélice, lo que da lugar a pequeños fragmentos, los fragmentos de
Okazaki, que forman la hebra retardada.
Para copiar cualquiera de las dos hebras se necesita la ADNpolimerasa III,
pero como ya se ha dicho, requiere que primero actúe una ARN polimerasa primer,
que ponga una secuencia de nucleotidos (de ARN) sobre los que se apoye la
ADNpolimrasa III. Una vez que se ha copiado la secuencia de la cadena de
ADN interviene una ADNpolimerasa I, que sustituye los fragmentos de ARN del cebador
por nucleotidos de ADN. Además, en el caso de la hebra retardada es necesario
la unión de los distintos fragmentos de Okazaki, para ello ha de actuar una ligasa.
A medida que se va realizando todo el proceso de copia y sustitución de los
cebadores, las ADNpolimerasa I, II presentan una actividad correctora de
errores, con capacidad endonucleasa
(reconocimiento del error y corte de la secuencia) y exonucleasa (eliminación de la zona erronea) y es sustituida por
los nucleótidos correctos por la ADNpolimerasa III.
Para reconocer la cadena de nueva síntesis
y la original ésta aparece metilada, proceso que se realiza tras la corrección
de errores.
vídeos: Horquilla de replicación
Replicación
Replicación

En el caso de Eucariotas
Los procesos de replicación del ADN en las células eucariotas son
semejantes, solo hay pequeñas diferencias:
-
presentan
un número mayor de replicones.
-
Existe
un número mayor de ADN polimerasas (cinco, α, β, γ, δ, ε)
- También
es necesario copiar las histonas, las histonas antiguas se quedan en la hebra
conductora, mientras que las nuevas histonas se incorporan a la hebra
retardada.
- Al
tratarse de un cromosoma lineal (el procariota era circular con lo que no
existen extremos), los extremos no pueden ser copiados, ya que al sustituirse
el cebador por ADN la ADN polimerasa debería “apoyarse por fuera de la cadena”,
de modo que la cadena se va acortando. Estos extremos constituyen los telómeros
del cromosoma, que presentan secuencias repetidas para precisamente evitar la
pérdida de información tras las sucesivas divisiones. No obstante, tras un
número determinado de divisiones, y por consiguiente la pérdida de determinado
número de copias de los telómeros, la célula entra en apoptosis.

Vídeo: Adición nucleótotidos durante la replicación


ANTERIOR apuntes pdf SIGUIENTE
DOGMA CENTRAL DE LA BIOLOGÍA MOLECULAR: REPLICACIÓN, TRANSCRIPCIÓN, TRADUCCIÓN-1/4


GENÉTICA MOLECULAR
ADN- Descubrimiento de la
molécula portadora de la información genética
La molécula que contenga la información debe ser estable, pero a la vez
debe tener cierta capacidad para el cambio, para poder asegurar la evolución,
no como cambio intencional, sino como cambio necesario para la supervivencia
ante modificaciones del ambiente. Además debe poder copiarse (replicarse) para
poder transmitir la información a la descendencia.
Ante estas características se proponían dos posibles moléculas: ácidos
nucleícos y proteínas. Y durante mucho tiempo se consideró que las proteínas
eran las que permitían las combinaciones y variabilidad suficiente, y fueron
consideradas las moléculas portadoras de la información.
En 1928 Griffith realiza experimentos con las bacterias de la neumonía (la
cepa encapsulada produce la muerte, mientras que las que no tienen cápsula no):
1- Griffith le inyecta a un ratón la cepa
encapsulada, el ratón muere
2- Inyecta la cepa sin cápsula a ratones y no
mueren
3- Por calor mata las bacterias de la cepa
virulenta y las inyecta a ratones, no mueren.
4- Prepara una mezcla de cepas virulentas
muertas por calor con cepas no virulentas pero vivas y las inyecta al ratón.
Los ratones mueren. Al aislar las bacterias causanten de la muerte observa que estas
bacterias presentan cápsula, es decir, de alguna manera la cepa no virulenta
sin cápsula se había “transfomado” en virulenta, desarrollando la cápsula.
Griffith propone la existencia de un
“factor transformante” que es capaz de transferirse entre organismos.
En 1944, Avery, McCarty y MacLeod
afirman que dicho factor transformante es el ADN.
Finalmente, en 1952 Hersey y Chase
demostraron que efectivamente la información genética está contenida en el ADN
y no en las proteínas. Para ellos realizaron un experimento con bacteriófagos y
bacterias.
1- Se prepararon dos grupos de bacteriófagos,
unos con las proteínas de la cápsida marcadas con S radioactivo, y otro grupo
con el ADN marcado con P radioactivo.
2- Se infectaron 2 grupos distintos de
bacterias con cada uno de los grupos de fagos.
3- Tras en proceso infeccioso observaron que
aquellos fagos que llevaban marcadas las proteínas, éstas se observaban fuera
de la célula, mientras que aquellos que llevaban marcado el ADN éste se
observaba en el interior celular. De este modo concluyeron que para que los
virus pudieran hacer copias de sí mismos tenían que inyectar el material que
tuviera la información para generar nuevos individuos, de modo que ese material
debía, obligatoriamente entrar al interior de la bacteria. Al observar que las
proteínas quedaban fuera, pero que el ADN entraba llegaron a la conclusión de
el material portador de la información para las nuevas generaciones era el ADN.
A todos estos estudios hay que añadir los de Chargaff en cuanto a la
proporción de los distintos componentes del ADN, y posteriormente 1953, la
estructura del ADN propuesta por Watson y Crick.
Antes de esto, Beadle y Tatum (1948)
demostraron la relación entre el ADN y las proteínas, a través de la inducción
de mutaciones en los ácidos nucleicos, lo que daba lugar a modificaciones en
las proteínas o incluso hacer que algunas llegaran a desaparecer.
Llegan a la conclusión de que un gen
codifica para una enzima, postulado que posteriormente se extiende a todos
los tipos proteicos. Tras esto se llega a la conclusión de que también puede
codificar para una cadena polipeptídica.

Con toda esta información, en 1970 Crick propone el Dogma Central de la Biología Molecular:
Es decir, el ADN daría siempre lugar a una copia en forma de ARN, proceso
de transcripción, el cual a su vez
daría lugar, por traducción, a las
proteínas. Por otra parte el ADN sería capaz de copiarse a sí mismo para
generar las copias para la descendencia, replicación.
Todos estos procesos requerirían de enzimas específicas para cada reacción.
No obstante, con el tiempo hubo que modificar este dogma, al descubrir que
el ARN también era capaz de copiarse a sí mismo. Y que además había organismos
(virus) cuya información genética era ARN capaz de generar una copia en forma
de ADN, en lugar de seguir la dirección para formar proteínas. A este nuevo
proceso se le llamó transcripción inversa o retotranscripción, llevada a cabo por la transcriptasa inversa.
SIGUIENTE apuntes pdf
domingo, 17 de abril de 2016
Suscribirse a:
Entradas (Atom)